sacharidy se tvoří v zelených rostlinách fotosyntézou, což je chemická kombinace nebo“ fixace “ oxidu uhličitého a vody využitím energie z absorpce viditelného světla. Celkovým výsledkem je snížení oxidu uhličitého na sacharidy a tvorba kyslíku:
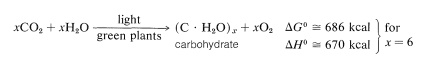
pokud je vytvořeným uhlohydrátem celulóza, pak je reakce ve skutečnosti opačná než spalování dřeva a zjevně vyžaduje značný přísun energie.
vzhledem k jeho životně důležitému charakteru pro život, jak ho známe, byla fotosyntéza intenzivně zkoumána a obecné rysy procesu jsou nyní poměrně dobře pochopeny. Mezi hlavní nedostatky v našich znalostech patří to, jak se světlo absorbované rostlinami přeměňuje na chemickou energii, a podrobnosti o tom, jak probíhá mnoho komplexních enzymem indukovaných reakcí.
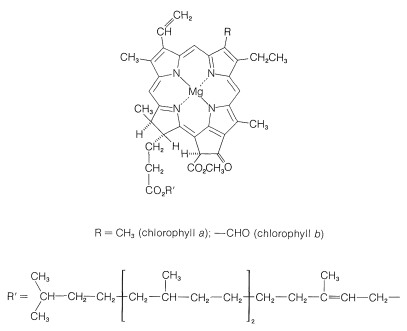
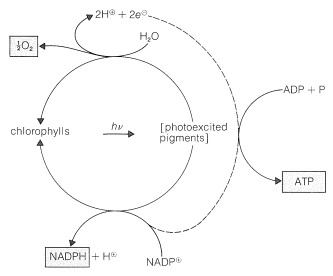
složky v zelených rostlinách, které vykonávají práci fotosyntézy, jsou obsaženy ve vysoce organizovaných membránově pokrytých jednotkách zvaných chloroplasty. Specifické látky, které absorbují světlo, jsou rostlinné pigmenty, chlorofyl a a chlorofyl b, jejichž struktury jsou znázorněny na obrázku 20-6. Tyto vysoce konjugované látky jsou velmi účinnými absorbéry světla a takto získaná energie se používá ve dvou samostatných procesech, které jsou znázorněny schematicky na obrázku 20-7.
jeden fotoproces redukuje nikotinamidadenin dinukleotid fosfát \(\left (\ce{NADP}^\oplus \ right)\) na \(\ce{NADPH}\). Tyto dinukleotidy, uvedené níže, se liší od \(\ce{NAD}^\oplus\) a \(\ce{NADH}\) (sekce 15-6C)tím, že mají fosfátovou skupinu na \(\ce{C_2}\) jedné z jednotek ribózy. Oxidovaná forma, \(\ce{NADP}^\oplus\), se chová jako \(\ce{NAD}^\oplus\) a přijímá ekvivalent \(\ce{H}^\ominus\) na \(\ce{C_4}\) nikotinamidového kruhu za vzniku \(\ce{NADPH}\):
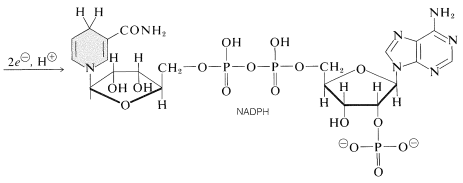
další důležitou fotoreakcí je oxidace vody na kyslík reakcí:
\
vytvořený kyslík jasně pochází z \(\ce{H_2O}\) a ne z \(\ce{CO_2}\), protože fotosyntéza v přítomnosti vody označené \(\ce{^{18}o}\) produkuje kyslík značený \(\ce{^{18}O}\), zatímco oxid uhličitý značený \(\ce{^{18}o}\) nedává kyslík značený \(\ce{^{18}o}\). Všimněte si, že oxidace vody produkuje dva elektrony a že tvorba \(\ce{NADPH}\) z \(\ce{NADP}^\oplus\) vyžaduje dva elektrony. Tyto reakce se vyskytují na různých místech v chloroplastech a v procesu přenosu elektronů z místa oxidace vody do místa redukce \(\ce{NADP}^\oplus\) se adenosin difosfát (ADP) převede na adenosintrifosfát (ATP; viz bod 15-5F pro diskusi o významu takových fosforylací). Transport elektronů mezi oběma fotoprocesy je tedy spojen s fosforylací. Tento proces se nazývá fotofosforylace (obrázek 20-7).
konečným výsledkem fotochemické části fotosyntézy je tvorba \(\ce{O_2}\), \(\ce{NADPH}\) a ATP. Většina kyslíku se uvolňuje do atmosféry, ale \(\ce{NADPH}\) a ATP se používají v řadě tmavých reakcí, které dosahují redukce oxidu uhličitého na hladinu a sacharidu (fruktózy). Vyvážená rovnice je
\
cyklus reakcí, které přeměňují oxid uhličitý na sacharidy, se nazývá Calvinův cyklus, po M. Calvin, který obdržel Nobelovu cenu za chemii v roce 1961 za svou práci na určování dráhy uhlíku ve fotosyntéze.
uhlík vstupuje do cyklu jako oxid uhličitý. Klíčová reakce, při které je \(\ce{CO_2}\) „fixována“, zahrnuje enzymatickou karboxylaci pentózy, \(D\) – ribulózy 1,5-fosfátu.\(^8\)
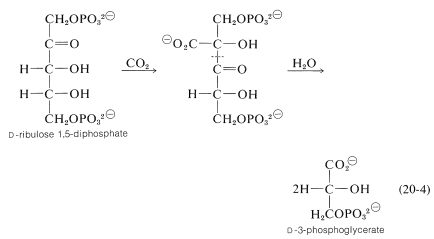
následným hydrolytickým štěpením \(\ce{C_2}\) – \(\ce{C_3}\) vazby karboxylačního produktu (to představuje reverzní Claisenovu kondenzaci; sekce 18-8B) se získají dvě molekuly \(D\)-3-fosfoglycerátu.\(^9\)
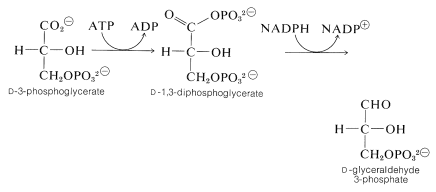
v následujících krocích se ATP používá k fosforylování karboxylové skupiny 3-fosfoglycerátu za vzniku 1,3-difosfoglycerátu(směsný anhydrid glycerinové a fosforečné kyseliny). Tato látka se pak redukuje o \(\ce{NADPH}\) na glyceraldehyd 3-fosfát:
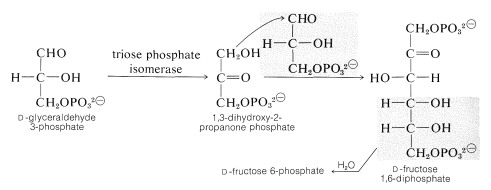
dva glyceraldehyd 3-fosfáty se používají k vytvoření šestiuhlíkového řetězce fruktózy aldolovou kondenzací \(\left (\ce{C_3} + \ce{C_3} \rightarrow \ce{C_6} \right)\), ale donorovým nukleofilem v této reakci je fosfátový ester dihydroxypropanonu, který je izomerem glyceraldehyd 3-fosfátu. Přeskupení \(\ce{C_3}\) aldózy na \(\ce{C_3}\) ketózu (typu popsaného v oddíle 20-2D)proto předchází přidání aldolu. (Pro diskusi o mechanismu enzymatické aldolové reakce viz bod 17-3F.) Vzniklý 1,6-difosfát fruktózy se pak hydrolyzuje na 6-fosfát fruktózy:
z toho, co jsme dosud popsali, byl z atmosféry přidán pouze jeden atom uhlíku, a přestože jsme dosáhli fruktózy, v tomto procesu bylo spotřebováno pět dříve redukovaných uhlíků. Rostlina tak musí získat zpět pětiuhlíkový cukr ze šestiuhlíkového cukru, aby udržela cyklus. Spíše než oddělit jeden uhlík a použít jej jako stavební blok ke konstrukci jiných cukrů, provádí se úžasná řada transformací, které lze shrnout pomocí následujících rovnic:
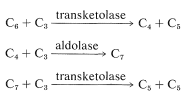
tyto reakce mají několik společných rysů. Všechny zahrnují fosfátové estery aldóz nebo ketóz a připomínají aldolové nebo reverzní aldolové kondenzace. Jejich mechanismy zde nebudou brány v úvahu, ale jsou podrobněji diskutovány v oddílech 20-10A, 20-10B a 25-10. Jejich součet je \(\ce{C_6} + 3 \ce{C_3} \rightarrow 3 \ ce{C_5}\), což znamená, že 6-fosfát fruktózy jako \(\ce{C_6}\) složka reaguje s celkem třemi \(\ce{C_3}\) jednotkami (dvěma glyceraldehyd 3-fosfáty a jedním dihydroxypropanon fosfátem) za vzniku tří ribulózových 5-fosfátů. I když se sekvence může zdát složitá, vyhýbá se vytváření pentózových nebo hexózových řetězců jeden uhlík najednou z meziproduktů jednoho uhlíku.
Calvinův cyklus je ukončen fosforylací \(D\) – ribulózy 5-fosfátu s ATP. Výsledný \(D\) – 1,5-difosfát ribulózy se pak použije k opětovnému zahájení cyklu kombinací s oxidem uhličitým. Existuje jedna šestina více fruktóza se používá k vytvoření dalších sacharidů, zejména glukózy, škrobu a celulózy.
\(^8\)Všechny reakce, o kterých budeme diskutovat, jsou zprostředkovány enzymy a od této chvíle vynecháme výslovnou zmínku o této skutečnosti. Nemělo by se však zapomínat, že se jedná o všechny procesy indukované enzymy, pro které máme jen málo, pokud vůbec, laboratorních činidel, která by se mohla duplikovat na konkrétních sloučeninách.
\(^9\)od této chvíle budeme v rovnicích označovat různé kyseliny, se kterými se setkáváme, jako fosfátové a karboxylátové anionty, i když to je sotva rozumné při hodnotách pH normálních v živých buňkách. Kyselina glycerová a kyselina fosforečná jsou pouze částečně ionizovány při pH 7-8. Bylo by však stejně nereálné reprezentovat kyseliny jako zcela nedisociované.
přispěvatelé a atributy
John D. Robert a Marjorie C. Caserio (1977) základní principy organické chemie, druhé vydání. W. A. Benjamin, Inc. , Menlo Park, CA. ISBN 0-8053-8329-8. Tento obsah je chráněn autorskými právy za následujících podmínek: „máte povolení k individuální, vzdělávací, výzkumné a nekomerční reprodukci, distribuci, zobrazení a provedení tohoto díla v jakémkoli formátu.“