kolhydrater bildas i gröna växter genom fotosyntes, som är den kemiska kombinationen, eller ”fixering”, av koldioxid och vatten genom utnyttjande av energi från absorption av synligt ljus. Det övergripande resultatet är minskningen av koldioxid till kolhydrat och bildandet av syre:
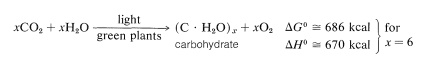
om det bildade kolhydratet är cellulosa, är reaktionen i effekt motsatt av förbränning av trä och kräver uppenbarligen betydande energiinmatning.
på grund av dess vitala karaktär för livet som vi känner det har fotosyntesen undersökts intensivt och processens allmänna egenskaper är nu ganska väl förstådda. De viktigaste bristerna i vår kunskap är hur ljuset som absorberas av växterna omvandlas till kemisk energi och detaljerna om hur de många komplexa enzyminducerade reaktionerna som är involverade äger rum.
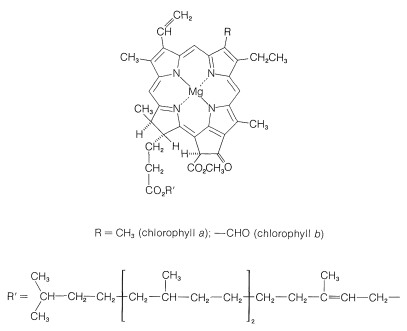
ingredienserna i gröna växter som utför fotosyntesens arbete finns i högorganiserade, membranbelagda enheter som kallas kloroplaster. De specifika ämnen som absorberar ljuset är växtpigmenten, klorofyll a och klorofyll b, vars strukturer visas i Figur 20-6. Dessa mycket konjugerade ämnen är mycket effektiva ljusabsorbenter, och den energi som erhålls används i två separata processer, som representeras schematiskt i Figur 20-7.
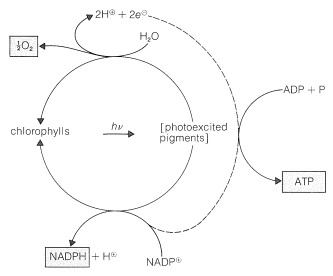
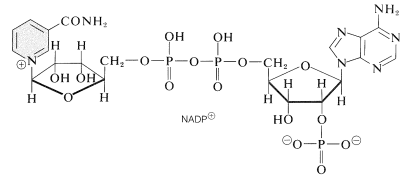
en fotoprocess reducerar nikotinamidadenindinukleotidfosfat \(\vänster (\ce{NADP}^\oplus\ höger)\) till \(\ce{NADPH}\). Dessa dinukleotider, som visas nedan, skiljer sig från \(\ce{nad}^\ oplus\) och \(\ce{NADH}\) (avsnitt 15-6c) genom att ha en fosfatgrupp vid \(\ce{C_2}\) hos en av ribosenheterna. Den oxiderade formen, \(\ce{NADP}^\oplus\), beter sig som \(\ce{NAD}^ \ oplus\) och mottar ekvivalenten av \(\ce{H}^\ ominus\) vid \(\ce{C_4}\) av nikotinamidringen för att bilda \ (\ce{NADPH}\):
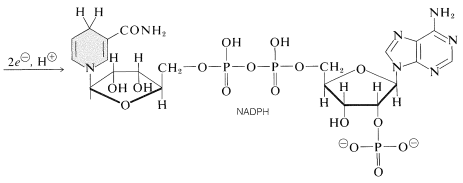
den andra viktiga fotoreaktionen är oxidation av vatten till syre genom reaktionen:
\
det syre som bildas kommer tydligt från \(\ce{H_2O}\) och inte från \(\ce{CO_2}\), eftersom fotosyntes i närvaro av vatten märkt med \(\ce{^{18}o}\) producerar syre märkt med \(\ce{^{18}O}\), medan koldioxid märkt med \(\ce{^{18}O}\) inte ger syre märkt med \(\ce{^{18}O}\). Observera att oxidationen av vattnet producerar två elektroner, och att bildandet av \(\ce{NADPH}\) från \(\ce{NADP}^\oplus\) kräver två elektroner. Dessa reaktioner uppträder på olika platser inom kloroplasterna och i processen att överföra elektroner från vattenoxidationsstället till reduktionsstället \(\ce{NADP}^\oplus\) omvandlas adenosindifosfat (ADP) till adenosintrifosfat (ATP; se Avsnitt 15-5F för diskussion mellan vikten av sådana fosforyleringar). Således är elektrontransport mellan de två fotoprocesserna kopplad till fosforylering. Denna process kallas fotofosforylering (figur 20-7).
slutresultatet av den fotokemiska delen av fotosyntesen är bildandet av \(\ce{O_2}\), \(\ce{NADPH}\) och ATP. Mycket av syret släpps ut i atmosfären, men \(\ce{NADPH}\) och ATP används i en serie mörka reaktioner som uppnår minskningen av koldioxid till nivån av A A kolhydrat (fruktos). En balanserad ekvation är
\
reaktionscykeln som omvandlar koldioxid till kolhydrater kallas Calvin-cykeln, efter M. Calvin, som fick Nobelpriset i kemi 1961 för sitt arbete med att bestämma vägen för kol i fotosyntes.
kol går in i cykeln som koldioxid. Nyckelreaktionen genom vilken \(\ce{CO_2}\) är ”fixerad” innefattar enzymatisk karboxylering av en pentos,\(D\)-ribulos 1,5-fosfat.\(^8\)
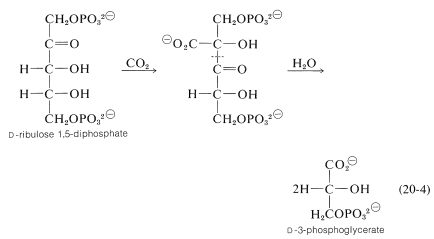
en efterföljande hydrolytisk klyvning av bindningen \(\ce{C_2}\)-\(\ce{C_3}\) av karboxyleringsprodukten (detta uppgår till en omvänd Claisen-kondensation; avsnitt 18-8b) ger två molekyler av \(D\)-3-fosfoglycerat.\(^9\)
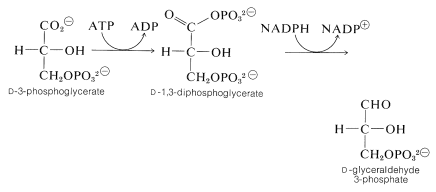
i efterföljande steg används ATP för att fosforylera karboxylgruppen av 3-fosfoglycerat för att skapa 1,3-difosfoglycerat (en blandad anhydrid av glycersyra och fosforsyror). Detta ämne reduceras sedan med \(\ce{NADPH}\) till glyceraldehyd 3-fosfat:
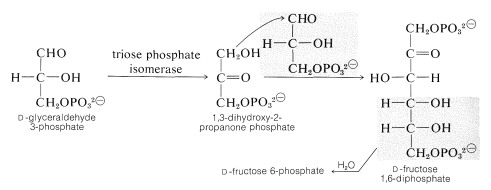
två glyceraldehyd 3-fosfater används för att bygga sexkolkedjan av fruktos genom en aldolkondensation \(\left (\ce{C_3} + \ce{C_3} \rightarrow \ce{C_6}\ right)\), men donatornukleofilen i denna reaktion är fosfatestern av dihydroxipropanon, som är en isomer av glyceraldehyd 3-fosfat. Omläggning av\ (\ce{C_3}\) Aldos till\ (\ce{C_3}\) ketos (av den typ som beskrivs i Avsnitt 20-2D) föregår därför aldoltillägget. (För en diskussion om mekanismen för den enzymatiska aldolreaktionen, se Avsnitt 17-3F.) Fruktos 1,6-difosfat bildas hydrolyseras sedan till fruktos 6-fosfat:
från vad vi hittills har beskrivit har endast en kolatom tillsatts från atmosfären, och även om vi har nått fruktos konsumerades fem tidigare reducerade kol i processen. Således måste växten få tillbaka ett femkolssocker från ett sexkolssocker för att fortsätta cykeln. I stället för att dela upp ett kol och använda det som en byggsten för att konstruera andra sockerarter, utförs en fantastisk serie transformationer som kan sammanfattas med följande ekvationer:
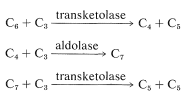
dessa reaktioner har flera funktioner gemensamt. De involverar alla fosfatestrar av aldoser eller ketoser, och de liknar aldol eller reverse-aldol kondensationer. Deras mekanismer kommer inte att övervägas här, men diskuteras mer uttryckligen i avsnitten 20-10a, 20-10b och 25-10. Deras summering är \(\ce{C_6} + 3 \ce{C_3} \rightarrow 3\ ce{C_5}\), vilket innebär att fruktos 6-fosfat som komponenten \(\ce{C_6}\) reagerar med totalt tre \(\ce{C_3}\) enheter (två glyceraldehyd 3-fosfater och ett dihydroxipropanonfosfat) för att slutligen ge tre ribulos 5-fosfater. Även om sekvensen kan verka komplex, undviker den att bygga upp pentos-eller hexoskedjor ett kol i taget från enkolsmellanprodukter.
Calvin-cykeln avslutas genom fosforylering av \(D\)-ribulos 5-fosfat med ATP. Den resulterande \(D\)-ribulos 1,5-difosfat används sedan för att starta cykeln igen genom att kombinera med koldioxid. Det finns en sjätte mer fruktos används för att bygga andra kolhydrater, särskilt glukos, stärkelse och cellulosa.
\(^8\)alla reaktioner vi kommer att diskutera förmedlas av enzymer, och vi kommer hädanefter att uttryckligen nämna detta faktum. Men det bör inte glömmas att det här är alla enzyminducerade processer, för vilka vi har få, om några, laboratoriereagenser att duplicera på de specifika föreningarna som är involverade.
\(^9\)Vi kommer hädanefter i ekvationer att beteckna de olika syrorna vi stöter på som fosfat-och karboxylatanjoner, även om detta knappast är rimligt vid pH-värdena normalt i levande celler. Glycer-och fosforsyror joniseras endast delvis vid pH 7-8. Det skulle emellertid vara lika orealistiskt att representera syrorna som helt oskadade.
bidragsgivare och attribut
John D. Robert och Marjorie C. Caserio (1977) grundläggande principer för organisk kemi, andra upplagan. W. A. Benjamin, Inc. , Menlo Park, CA. ISBN 0-8053-8329-8. Detta innehåll är upphovsrättsskyddat under följande villkor, ” du får tillstånd för individuell, pedagogisk, forskning och icke-kommersiell reproduktion, distribution, visning och utförande av detta arbete i vilket format som helst.”