炭水化物は、可視光の吸収からのエネルギーを利用して二酸化炭素と水の化学的組み合わせ、または”固定”である光合成によって緑の植物に形成される。 全体的な結果は、炭水化物への二酸化炭素の減少および酸素の形成である:
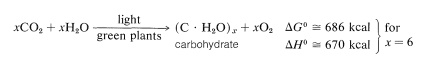
形成された炭水化物がセルロースである場合、実際の反応は木材の燃焼の逆であり、明らかにかなりのエネルギー入力を必要とする。
私たちが知っているように生命にとって重要な性質のために、光合成は集中的に調査されており、プロセスの一般的な特徴はかなりよく理解されて 私たちの知識の主な欠点は、植物によって吸収された光がどのように化学エネルギーに変換されるか、そして多くの複雑な酵素誘発反応がどのよう
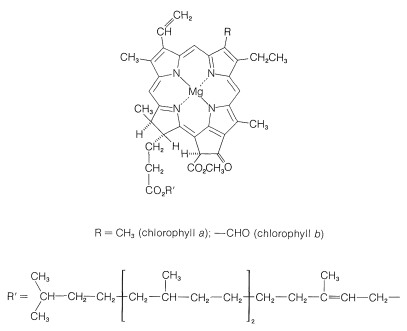
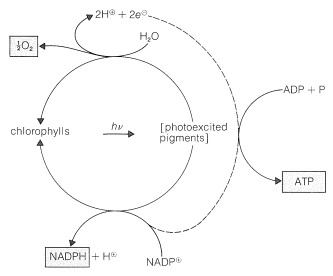
光合成の働きを続ける緑の植物の成分は、葉緑体と呼ばれる高度に組織化された膜で覆われた単位に含まれています。 光を吸収する特定の物質は、植物色素であるクロロフィルaとクロロフィルbであり、その構造は図20-6に示されています。 これらの高度に共役した物質は非常に効率的な光吸収体であり、そのように得られたエネルギーは2つの別々のプロセスで使用され、図20-7に図式的に表
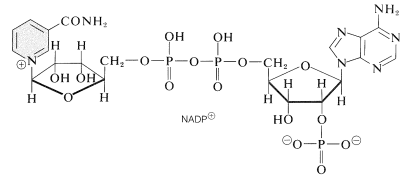
一つの光プロセスは、ニコチンアミドアデニンジヌクレオチドリン酸\(\left(\ce{NADP}^\oplus\right)\)を\(\ce{NADPH}\)に還元する。 以下に示すこれらのジヌクレオチドは、リボース単位の1つの\(\ce{C_2}\)にリン酸基を有する点で、\(\ce{NAD}^\oplus\)および\(\ce{NADH}\)(セクション15-6C)とは異なる。 酸化された形\(\ce{NADP}o\oplus\)は\(\ce{NAD}o\oplus\)のように振る舞い、ニコチンアミド環の\(\ce{C_4}\)で\(\ce{H}^\ominus\)と同等のものを受け取り、\(\ce{NADPH}form\oplus\)を形成する。}\):
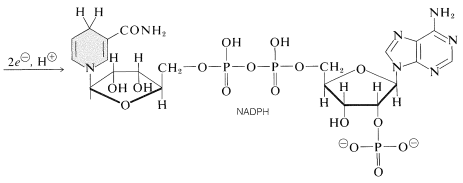
他の重要な光反応は、反応による水の酸素への酸化である:
\
これは、\(\ce{H{18}O}\)で標識された水の存在下での光合成は\(\ce{H{18}O}\)で標識された酸素を生成するのに対し、\(\ce{H{18}O}\)で標識された二酸化炭素は\(\ce{^{18}O}\)で標識された酸素を生成しないためです。 水の酸化は2つの電子を生成し、\(\ce{NADPH}from\oplus\)から\(\ce{NADPH}.\oplus\)の形成には2つの電子が必要であることに注意してください。 これらの反応は、葉緑体内の異なる位置で起こり、水酸化部位から\(\ce{NADP}^\oplus\)還元部位に電子を移動させる過程で、アデノシン二リン酸(ADP)はアデノシン三リン酸(ATP;そのようなリン酸化の重要性の間の議論についてはセクション15-5Fを参照)に変換される。 したがって、二つの光プロセス間の電子輸送は、リン酸化に結合される。 このプロセスは光リン酸化と呼ばれます(図20-7)。光合成の光化学部分の最終結果は、\(\ce{O_2}\)、\(\ce{NADPH}\)、およびATPの形成である。 酸素の多くは大気中に放出されますが、\(\ce{NADPH}\)とATPは、二酸化炭素をa炭水化物(フルクトース)のレベルまで減少させる一連の暗反応に利用されます。\(\ce{NADPH}\)とatpは、\(\ce{NADPH}\)とatpは、\(\ce{NADPH}\)とatpは、\( 平衡方程式は次のようになります
\
二酸化炭素を炭水化物に変換する反応のサイクルは、Mの後にカルビンサイクルと呼ばれています。 カルビンは、光合成における炭素の経路を決定する彼の仕事のために1961年にノーベル化学賞を受賞しました。
炭素は二酸化炭素としてサイクルに入ります。 \(\Ce{CO_2}\)を\(\ce{CO_2}\)で固定する主な反応には、ペントース\(D\)-リブロース1,5-リン酸の酵素的カルボキシル化が含まれる。\(^8\)
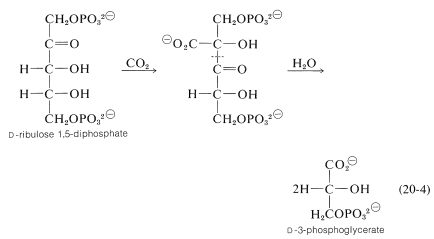
カルボキシル化生成物の\(\ce{C_2}\)-\(\ce{C_3}\)結合のその後の加水分解切断(これは逆クライゼン縮合になる;セクション18-8B)は、\(D\)-3-ホスホグリセリン酸の二つの分子をもたらす。\(^9\)
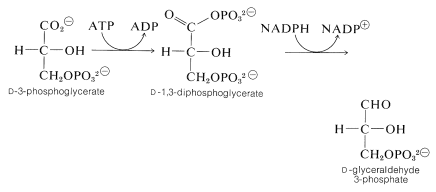
それに続くステップでは、ATPが1,3-diphosphoglycerate(グリセリックおよびリン酸の混合された無水物)を作成するために3-phosphoglycerateのカルボキシル基をリン酸化するのに利 この物質はその後、\(\ce{NADPH}\)によってグリセルアルデヒド3-リン酸に還元されます:
2つのグリセルアルデヒド3-リン酸塩は、アルドール縮合によってフルクトースの6炭素鎖を構築するために利用されている\(\left(\ce{C_3}+\ce{C_3}\rightarrow\ce{C_6}\right)\)が、この反応におけるドナー求核剤は、グリセルアルデヒド3-リン酸の異性体であるジヒドロキシプロパノンのリン酸エステルである。 したがって、\(\ce{C_3}\)アルドースを\(\ce{C_3}\)ケトース(セクション20-2Dで説明されているタイプ)に再配置すると、アルドール添加が先行します。 (酵素的アルドール反応の機構の議論については、セクション17-3Fを参照してください。)形成されたフルクトース1,6-二リン酸は、次いでフルクトース6-リン酸に加水分解される:
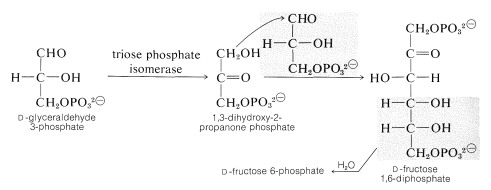
これまでに説明したことから、大気から炭素の原子が1つだけ添加されており、フルクトースに達していますが、以前に還元された炭素が5つ消費され したがって、植物はサイクルを永続させるために六炭素砂糖から五炭素砂糖を取り戻す必要があります。 一つの炭素を分割し、他の糖を構築するためのビルディングブロックとしてそれを使用するのではなく、次の式で要約することができる驚くべき一連:
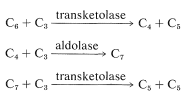
これらの反応にはいくつかの共通点があります。 それらはすべてアルドースまたはケトースのリン酸エステルを含み、アルドールまたは逆アルドール縮合に似ている。 それらのメカニズムはここでは考慮されませんが、セクション20-10A、20-10B、および25-10でより明示的に議論されています。 それらの合計は\(\ce{C_6}+3\ce{C_3}\rightarrow3\ce{C_5}\)であり、これは\(\ce{C_6}\)成分としてのフルクトース6-リン酸塩が合計3つの\(\ce{C_3}\)単位(2つのグリセルアルデヒド3-リン酸塩と1つのジヒドロキシプロパノンリン酸塩)と反応して最終的に3つのリブロース5-リン酸塩を与えることを意味する。 配列は複雑に見えるかもしれないが、それは一炭素中間体から一炭素ずつペントース鎖またはヘキソース鎖を構築することを回避する。
カルビンサイクルは、ATPによる\(D\)-リブロース5-リン酸のリン酸化によって完了する。 得られた\(D\)-リブロース1,5-二リン酸は、二酸化炭素と組み合わせることによって再びサイクルを開始するために使用されます。 他の炭水化物、特にグルコース、デンプン、セルロースを構築するために使用される六分の一以上のフルクトースがあります。
\(^8\)我々が議論するすべての反応は酵素によって仲介されており、この事実についての明示的な言及は今後省略する。 しかし、これらはすべて酵素誘導プロセスであることを忘れてはならず、関与する特定の化合物に複製する実験室試薬があれば、ほとんどありません。
\(^9\)これは生きている細胞の正常なpH値ではほとんど合理的ではありませんが、我々は今後、式では、我々が遭遇する様々な酸をリン酸塩およびカルボキシレートアニオンとして指定します。 グリセリックおよびリン酸は、pH7-8で部分的にのみイオン化される。 しかし、酸を完全に解離していないものとして表現することは同様に非現実的であろう。
貢献者と帰属
John D.Robert and Marjorie C.Caserio(1977)Basic Principles of Organic Chemistry,second edition. W.A.Benjamin,Inc. 、メンロパーク、カリフォルニア州。 ISBN0-8053-8329-8. このコンテンツは、以下の条件の下で著作権で保護されています,”あなたは、個人のための許可を与えられています,教育,研究および非商業的な複製,配”